Generating More Robustly Infected Cell Lines by Co-Culture Infection

This article was contributed by:
Matthew Gardner
Graduate Student, Glaunsinger LabUniversity of California, Berkeley
Viral Infection in Tissue Culture
Herpesviruses are large, double-stranded DNA viruses that have evolved over millions of years to become master manipulators of cellular machinery. By studying how these viruses—and viruses in general—hijack host cell machinery, we can learn a great deal not only about strategies of viral pathogenesis, but also about the fundamental cellular systems involved.
Given the highly-specialized nature of these viruses, it is difficult to mimic a natural infection in a tissue culture setting. Even when viruses can be cultured and studied, genomic mutants are often difficult to produce or cannot be maintained in the tissue culture system optimized for the wild-type virus. This was the case for Kaposi’s sarcoma-associated herpesvirus (KSHV), which we study in the Glaunsinger Lab. Finding a way to stably infect cell lines with mutant KSHV took the entire field many years of work.
The Stability Problem
The first major hurdle in the manipulation of mutant KSHV was cleared in 2012, when robust genetic tools for KSHV were developed.1 The system uses a bacterial artificial chromosome (BAC) that harbors the KSHV genome. This BAC can be propagated in Escherichia coli and manipulated by the Red recombinase system present in the bacteria. There have been some minor improvements to the original protocol by our lab to improve mutagenesis—most notably, we have used gBlocks as the mutational insert. However, a major stumbling block remained: the generation of stably-infected cell lines.
Generating a stable infection is a common problem in the study of host-pathogen interactions. The pathogen of interest often outright kills the target cells, or the target cells detect and inactivate the pathogen. In the case of KSHV, the virus generally does not have difficulty evading the host response. But, depending on the method of infection, the virus may act inconsistently. For example, cell lines generated by direct transfection of the purified BAC or by infection with purified, cell-free virus remain infected, but they lose the ability to express viral genes or to produce new virions over time. Even when viral production is initiated, it is initiated in an attenuated state. When measuring viral genome replication by qPCR, we expected approximately 30-fold DNA replication, but we only observed 5 to 10-fold DNA replication. We also found that very few infectious particles were produced.
Natural Infections Provide a Solution
The key to generating a stable cell line turned out to be mimicking a more natural infection, where cells are rarely infected with cell-free virus and never by naked viral genomes. Dr. Zsolt Toth (University of Florida) and Dr. Jae Jung (University of Southern California) found that a more natural route of infection led to a better-behaved cell line. In the microenvironment of a Kaposi’s sarcoma tumor, only 1-2% of cells are in the virion-producing lytic phase at any time. The majority of cells are in the latent phase, expressing only a subset of the ~90 viral genes. The few genes that are expressed during latency are required to maintain the tumor microenvironment,2 which is densely-packed and highly-inflammatory. This environment is more closely mimicked by a co-culture infection model than by a cell-free virus infection model. In the Glaunsinger Lab, we perform co-culture infection according to the protocol outlined below, developed with guidance from Dr. Toth.
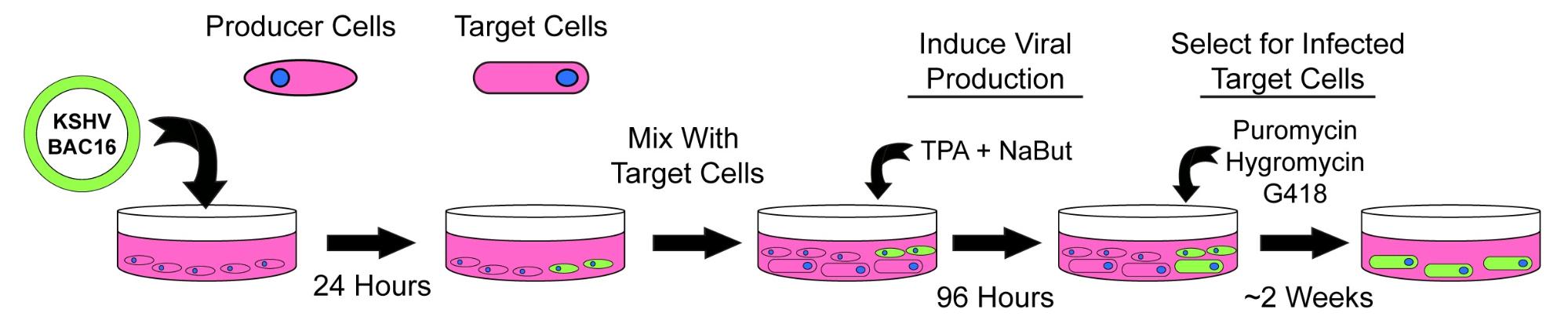
Figure 1. A method for mimicking a natural KSHV infection.
How to Perform a Co-Culture Infection
First, a BAC16 clone is produced using a slightly modified version of the Brulois protocol.1,3. The purified BAC is then transfected into HEK293T cells, which will serve as the producer cell line. HEK293T cells are used because they are easily transfected, fast-growing, and sensitive to selection drugs. After 24 hours, the producer cells are checked for infection; infected cells will constitutively express GFP, encoded by the viral genome.

Figure 2. Progression of KSHV infection during drug selection.
Producer cells (transfected HEK293T cells) are next co-cultured with target cells (uninfected iSLK cells). Both types of cells are trypsinized, counted, and mixed at a 1:1 ratio in a new vessel. The co-culture vessel should be large enough to comfortably accommodate the total number of cells that are added, but small enough to maintain them in close contact. Once the cells are mixed, viral production is induced by the addition of TPA (PMA) and sodium butyrate (NaBut). After four days, the cells should cover the entire tissue culture vessel, with iSLK puro cells growing as an even layer on the bottom and the HEK293T cells growing in clumps on top of them (Figure 2).
Infected target cells are isolated over the next two weeks through the addition of selection drugs, which kill the producer cells and uninfected target cells. The new cell line is then expanded and frozen in small aliquots for future studies. In the Glaunsinger lab, we generally characterize any new cell line by assessing viral gene expression, DNA replication, and infectious virion production.
For a detailed protocol, please see this protocol in Benchling.
Co-Culture Infection Enables the Discovery of KSHV ORF68's Role
Recently, we used this protocol to study the role of the KSHV protein ORF68. This protein had not previously been studied, although homologous proteins in related viruses suggested that it would play a role in viral DNA packaging. We showed that ORF68 was required for the production of infectious virions. While the wild-type virus could infect new cells, the mutant virus could not, despite replicating the viral genome. Furthermore, we showed that without ORF68, the viral genome is not cleaved, an event integral to the production of new virions. This lead us to purify and characterize ORF68 in vitro. We discovered that it was a DNA-binding protein associated with nuclease activity.3
Without the cell-based viral characterization enabled by co-culture infection, I doubt we would have uncovered this activity. ORF68 is of particular significance because it is an essential viral protein, required for virion production. This study demonstrated how we can use the co-culture infection protocol to study viruses with mutations in essential genes. The only modification required when working with lethal mutations is to use a HEK293T cell line that stably expresses the gene that you are mutating, which in our case was ORF68.
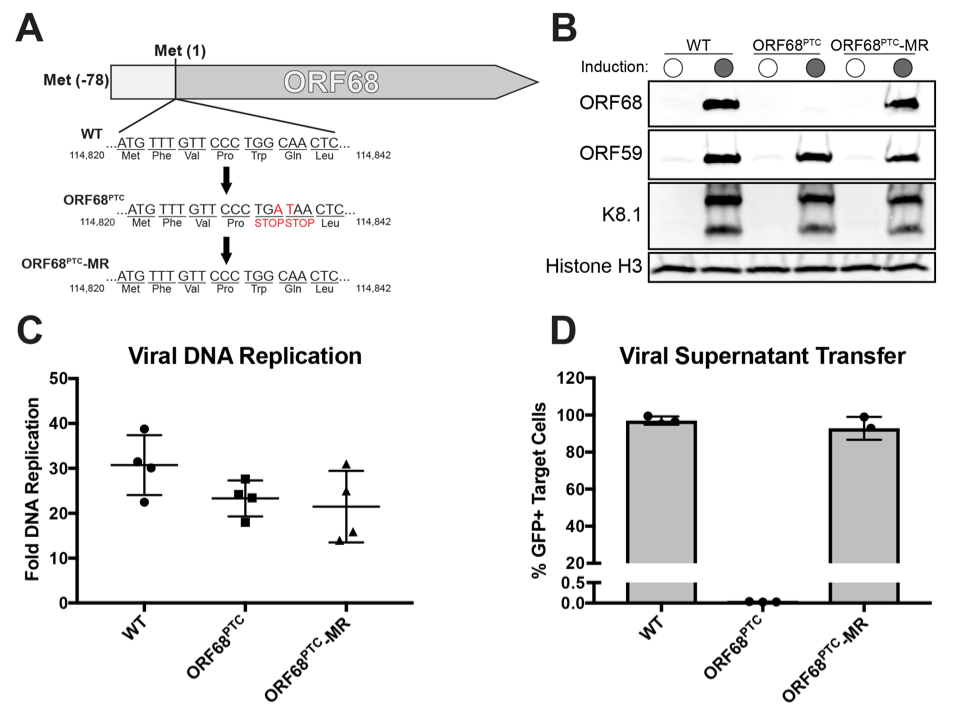
Figure 3. Characterization of the ORF68-deficient (ORF68PTC) mutant virus. (A) Diagram depicting the genetic locus of ORF68 and the mutations inserted into the ORF68PTC virus. (B) Western blots showing expression of the early ORF59 protein and the late K8.1 protein in WT, ORF68PTC, and ORF68PTC-MR viruses. Histone H3 served as a loading control. (C) DNA replication was measured by qPCR of the viral genome before and after induction of the lytic cycle. (D) Progeny virion production was assayed by supernatant transfer and flow cytometry of target cells.3
References
Editor's Note: The featured photo for this article is a TEM image, provided to us by the author. In this image, viral capsids—the dark, geometric shapes—are visible inside a host cell.
Matthew Gardner is an academic who uses Benchling. Learn more:
If you or your colleagues are interested in contributing an article about your research to the Benchling blog, let us know!